
The morphological features that unite all echinoderms are the water vascular system and a mesodermal skeleton comprised of numerous plates. Each plate is a single crystal of calcite. Calcite is a mineral that is readily preserved through geological time; however, the water vascular tissue is entirely soft tissue and not preserved. Fortunately, the track of the water vascular system can be preserved because an ambulacrum sits along a complex of distinctive plates. Thus, both of the criteria needed to define an echinoderm can be preserved in fossils.
The oldest definite echinoderms are from the lower Cambrian, more than 540 million years ago (Ma). Older multicellular echinoderms have been reported from Ediacaran strata (ca. 600 Ma), but specialists debate whether any of these fossils represent echinoderms or some other organism.
The phylum Echinodermata is subdivided into clades/classes that are generally differentiated by the geometry of the body, symmetry of the body, presence or absence of appendages, and the type of plating. Three basic body plans appeared among early Cambrian echinoderms, which served as the seed for the Cambrian echinoderm radiation. These three body plans are represented by
1) Ctenocystoids: lateralized and symmetrical/asymmetrical body plan, with or without appendages, and tessellate plating (Fig. 1)
2) Helicoplacoidea: ellipsoidal-shaped body, obvious helical symmetry, no appendages, and imbricate plating (Fig. 2); and
3) Edrioasteroid: disc-shaped body, pentaradiate symmetry, no appendages, and imbricate and tesselate plating (Fig. 3).

One of the primary objectives of the EchinoToL project is to decipher how younger echinoderm classes are rooted with these earliest, divergent body plans.
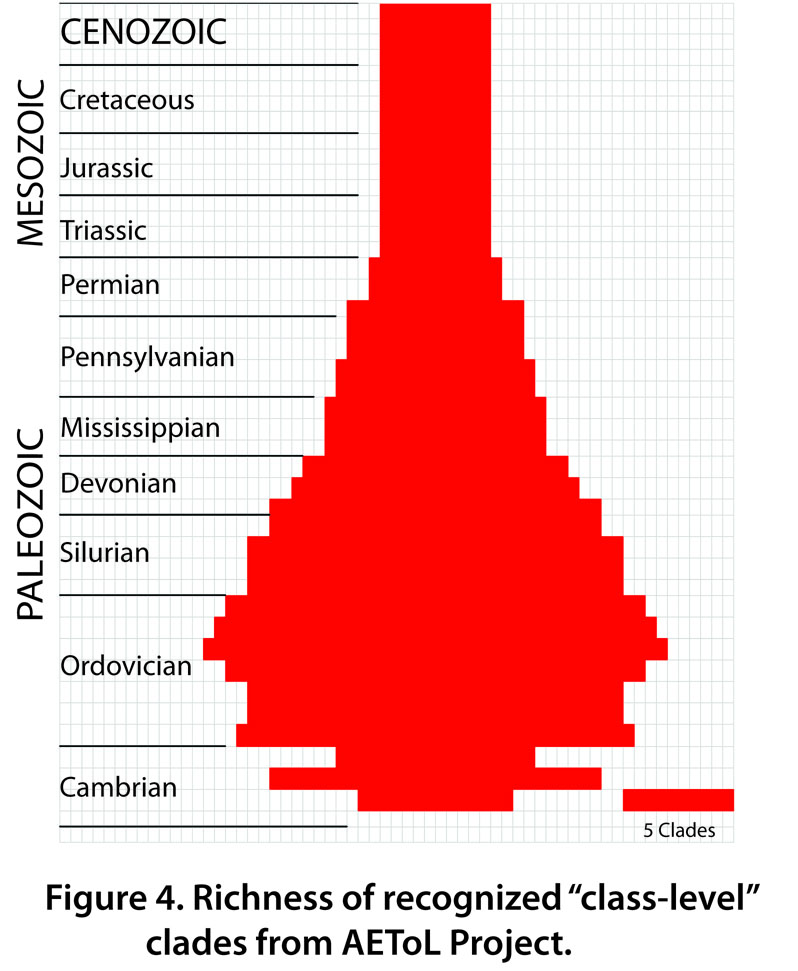 For the EchinoToL project, as many as 33 “class-level” clades are recognized among the Echinodermata (Fig. 4). Significant clade diversification occurred twice: from the early to the middle Cambrian and from the Middle to the Late Ordovician. Clade richness increased to 15 by the middle Cambrian (Fig. 4) largely due to the first appearance in the fossil record of lateralized echinoderm groups [stylophorans (Fig. 5), cinctans, solutes, and ctenocystoids (Fig. 6)], edrioasteroid groups, and erect pentaradiate groups [e.g., gogiids (Fig. 6), imbricates, and helicocystoids]. Diversity appears to have decreased into the late Cambrian, but this may be an artifact of preservation.
For the EchinoToL project, as many as 33 “class-level” clades are recognized among the Echinodermata (Fig. 4). Significant clade diversification occurred twice: from the early to the middle Cambrian and from the Middle to the Late Ordovician. Clade richness increased to 15 by the middle Cambrian (Fig. 4) largely due to the first appearance in the fossil record of lateralized echinoderm groups [stylophorans (Fig. 5), cinctans, solutes, and ctenocystoids (Fig. 6)], edrioasteroid groups, and erect pentaradiate groups [e.g., gogiids (Fig. 6), imbricates, and helicocystoids]. Diversity appears to have decreased into the late Cambrian, but this may be an artifact of preservation.

The most significant diversification of echinoderm clades occurred during the Great Ordovician Biodiversification Event (GOBE), when suspension-feeding organisms came to dominate ocean floor habitats. Richness of “class-level” clades during the Middle Ordovician was as high as 21 (Figure 4), including, among others, asteroids (Fig. 7), blastoids (Fig. 8), coronoids, crinoids (Fig. 9), cyclocystoids, diploporans (Fig. 10), echinoids (Fig. 11), edrioasteroids (Fig. 3), edrioblastoids, eocrinoids (Fig. 2), holothurians, homoiostelians (=solutes), ophiocistioids, ophiuroids (Fig. 12), parablastoids, paracrinoids, rhombiferans (Fig. 13), and stylophorans (Fig. 6). This diversity was the result of three Cambrian classes becoming extinct, six classes that survived from the Cambrian into the Ordovician, and eleven new classes that originated during the early and middle Ordovician.
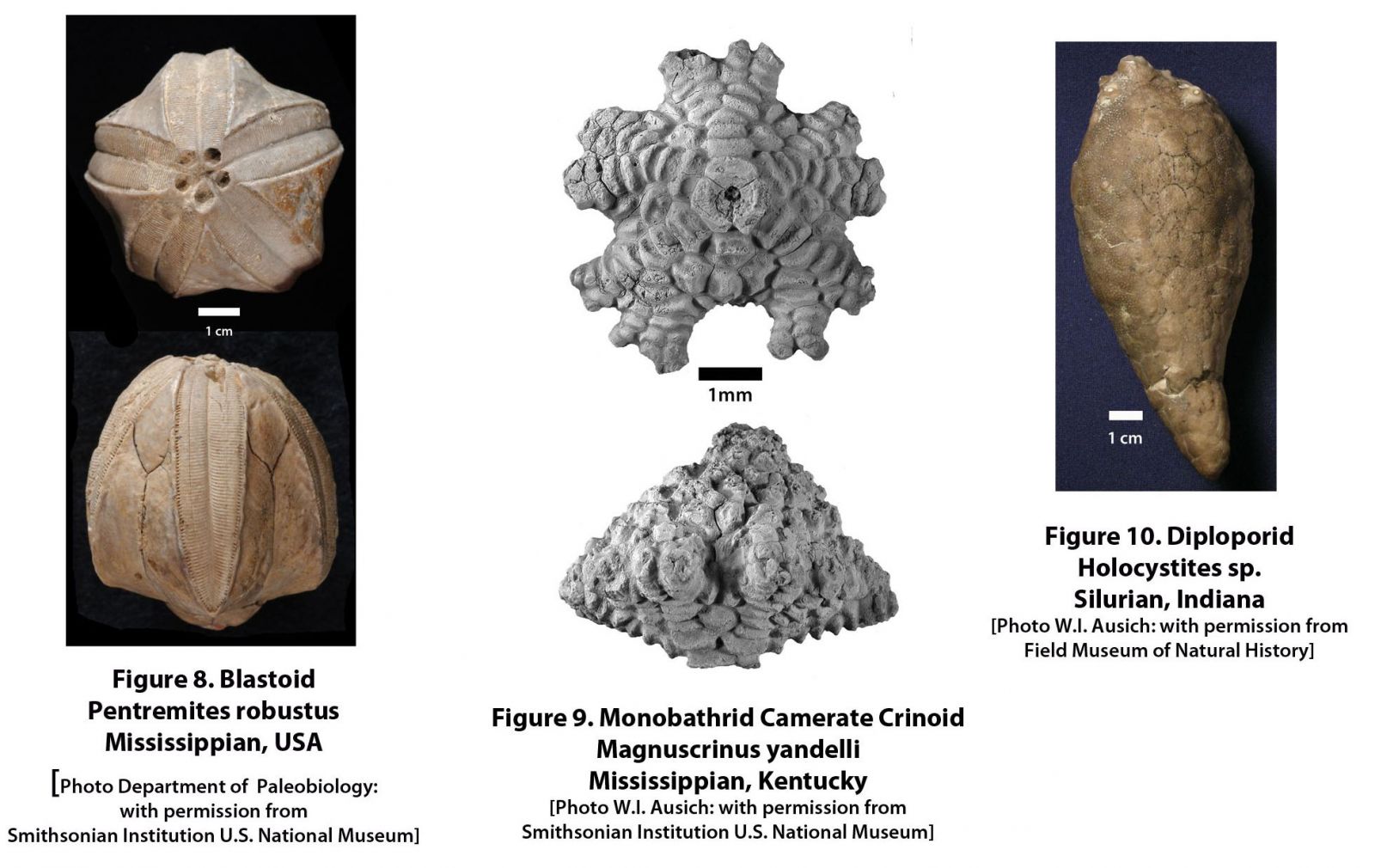
 From the Ordovician peak, echinoderm class-level richness gradually declined through the remainder of the Paleozoic, such that there were 17 echinoderm classes during the middle Silurian, 13 during the middle Devonian, 10 during the middle Mississippian, 9 during the middle Pennsylvanian, and 6 during the middle Permian. As echinoderm classes dwindled, the representation of crinoids increased. The only significant exception is that blastoids were locally very abundant in select environments during the middle and late Paleozoic.
From the Ordovician peak, echinoderm class-level richness gradually declined through the remainder of the Paleozoic, such that there were 17 echinoderm classes during the middle Silurian, 13 during the middle Devonian, 10 during the middle Mississippian, 9 during the middle Pennsylvanian, and 6 during the middle Permian. As echinoderm classes dwindled, the representation of crinoids increased. The only significant exception is that blastoids were locally very abundant in select environments during the middle and late Paleozoic.
With the extinction of blastoids, ophiocistiods, and isorophid edrioasteroids during the Permian, only five classes – asteroids, crinoids, echinoids, holothurians, and ophiuroids – survived into the Mesozoic (Fig. 4). Each of these five classes was or is dominant in certain environmental settings or at certain times during the past. However, echinoids have been the dominant post-Paleozoic echinoderm group, both in terms of number of specimens and number of genera and species (Fig. 14).
No new echinoderm classes have evolved since the Ordovician. The class-level richness in today’s oceans remains at five. Only the echinoderm classes that survived the great end-Permian mass extinction remain alive today.
Work during the past few years and continuing with the Echinoderm Tree of Life Project will refine the history described above. With more detailed phylogenetic studies, clades will be more rigorously defined, and “class-level” clade richness may increase further, especially during the Cambrian. Discoveries of new fossil echinoderm specimens, especially from the lower Paleozoic of the Gondwana paleocontinent (northern Africa), are yielding both new, bizarre clades and stratigraphic range extensions of clades not previously known from Cambrian strata.